
Cameraria ohridella Deschka & Dimić, 1986
szrotówek kasztanowcowiaczek
Horse Chestnut Leafminer |   | Rodzina | | Gracillariidae kibitnikowate | | Rząd | | Lepidoptera motyle | | Gromada | | Insecta owady | | Typ | | Arthropoda stawonogi | | |
 | |  | | | | | | | | |
 | | | | | | Locus typicus/Terra typica | | Okolice jez. Ochrydzkiego (Macedonia); gatunek opisany w stadium ekspansji. | | | | | | Pierwotna ojczyzna gatunku | | Pierwotna ojczyzna gatunku na razie nie jest znana. Są tu trzy możliwości: 1) nagłe uaktywnienie się nieznanego wcześniej (przeoczonego wcześniej przez entomologów?) drobnego motyla z Anatolii i południowego rejonu Bałkanów, gdyż tam w dolnym reglu (700-1200 m n.p.m.) występuje w domieszce preferowany przez niego kasztanowiec biały (Aesculus hippocastanum(L.); 2) zawleczenie szrotówka (np. drogą lotniczą) ze wschodniej części Ameryki Północnej, gdzie występuje kilka gatunków kasztanowców oraz żyją inne formy z rodzaju Cameraria; 3) zawleczenie transportowe motyli z południowej lub wschodniej Azji, gdzie również oba te rodzaje są reprezentowane. Przystosowania biologiczne tego motyla (diapauza zimowa w grubszym kokonie) wskazują na pochodzenie z obszaru klimatu umiarkowanego, a więc raczej na pierwszą lub drugą proweniencję. Jednak pierwsza możliwość wydaje się mniej prawdopodobna, gdyż kasztanowce białe wprowadzane były do parków pałacowych Europy począwszy od 1576 r. (cesarski ogród w Wiedniu), m.in. do królewskiego ogrodu w Krakowie-Łobzowie ok. 1580 r. i wobec niewielkich odległości od południowych Bałkanów szrotówek miałby dostateczną ilość czasu aby podążyć za żywicielem na północ, chociażby korzystając z częstych w tych obszarach, wiejących wiosną i latem, wiatrów z południa. Może jednak należy uwzględnić też wpływ skokowego ocieplenia klimatu w ostatnich dekadach XX wieku i powstania mutacji gradacyjnej wykorzystującej występowanie w Europie alej wysadzanych kasztanowcami i zwartych grup tych drzew w parkach, gdyż faktycznie powstały w ten sposób zwarte monokultury kasztanowca, który w obszarze naturalnego areału jest tylko gatunkiem domieszkowym. Teoretycznie obie te okoliczności mogłyby spowodować gradację szrotówka na Bałkanach i szybką jego ekspansję na północ; w takim przypadku byłaby to inwazja regionalna.
Pozostaje więc inwazja międzykontynentalna, za czym przemawia brak wcześniejszych stwierdzeń przedstawicieli Cameraria w Europie. Jednak ostateczne i jednoznaczne określenie obszaru wyjściowego opisywanej inwazji będzie możliwe dopiero po wnikliwej rewizji amerykańskich i azjatyckich form tego rodzaju - nie można bowiem wykluczyć przypadku, iż opisany niedawno z Macedonii inwader może okazać się jednym ze znanych już gatunków egzotycznych. | | | | | | Podstawowe cechy morfologiczne | | Drobny motyl o rozpiętości skrzydeł 2,8-3,8 mm, koloru złoto-ochrowego z poprzecznymi białymi plamami, z wąskim czarnym obrzeżeniem. Głowa z pióropuszem srebrzystych włosków, czułki nitkowate długości skrzydeł. | | | | | | Biologia, ekologia | | Sukcesy inwazyjne szrotówka zmobilizowały europejskie służby ochrony roślin do przyśpieszonych badań jego biologii i ekologii (Pschorn-Walcher 1997, Gregor i in. 1998, Skuhravý 1999, Svatos i in. 1999, Freise i Heitland 2001, Kosibowicz 2005). Motyl wyraźnie preferuje kasztanowca białego (Aesculus hippocastanum) oraz jego odmiany "Pyramidalis" i "Baumannii". Rozwija się także na kasztanowcu japońskim (Ae. turbinata ). W znacznie mniejszym stopniu uszkadza liście kasztanowca żółtego (Ae. flava), kasztanowca krwistego (Ae. pavia) i kasztanowca czerwonego (Ae. x carnea), nie rozwija się natomiast w liściach kasztanowca gładkiego (Ae. glabra) i kasztanowca drobnokwiatowego (Ae. parviflora, Ae. indica). Niekiedy w przypadku silnej gradacji samice nie mają możliwości złożenia jaj na kasztanowcach i wówczas jaja składane są na liściach drzew i krzewów rosnących w pobliżu, takich jak jesion (Fraxinus) buk (Fagus), tawuła (Spiraea) czy śnieguliczka (Symphoricarpus), jednakże larwy się na nich nie rozwijają.
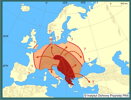
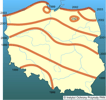
Obserwacje prowadzone w Polsce i krajach ościennych wykazały, że szrotówek może rozwijać się na klonach (Acer spp.). Szczególnie często obserwowano zasiedlanie jaworu (Acer pseudoplatanus) nawet w miejscach odległych od nasadzeń kasztanowców (Kosibowicz 2003). Na liściach jaworu rozwój szrotówka przebiega analogicznie jak na kasztanowcu, a wylatujące z min motyle nie różnią się morfologicznie od motyli wylatujących z liści kasztanowca. Rozwój na jaworach podawano już wcześniej z Austrii i Czech, jednak w krajach tych było to zjawisko marginalne.
Wiele ważnych informacji o biologii i ekologii szrotówka wniosły badania (2001). Dorosłe motyle latają słabo i na niewielkich dystansach, jednak z pomocą wiatru i transportu lądowego mogą przenosić się na duże odległości, co, w połączeniu z 3-4 pokoleniami w roku, ułatwia im opanowanie w krótkim czasie stosunkowo dużych przestrzeni (100-150 km rocznie). Motyle pojawiają się wiosną, w okresie, gdy kasztanowce pokryte są już liśćmi; ma to miejsce z reguły w końcu kwietnia. Wyloty rozciągnięte są w czasie, a ich kulminacja następuje w pierwszych dniach maja. Po wydostaniu się z kokonów znajdujących się w zeszłorocznych opadłych liściach - motyle nalatują na pnie kasztanowców. Na pniach niektórych drzew w tym okresie obserwowano setki przesiadujących motyli. Tu też dochodzi do kopulacji, po czym zapłodnione samice przystępują do składania na liściach jaj. Jedna samica składa ich około 40. Jaja wielkości 0,2-0,3 mm składane są zawsze na zewnętrznej stronie liścia w zagłębieniach w pobliżu nerwów głównych i bocznych liści. Kształt jaja jest soczewkowaty o chropowatej i nieregularnej powierzchni. W chwili złożenia jest ono przeźroczyste, a w miarę rozwoju embrionalnego staje się mlecznobiałe. Embrion rozwija się w jaju przez 10 do 20 dni w zależności od warunków atmosferycznych. Larwa bezpośrednio z jaja dostaje się do liścia i w pierwszym okresie rozwoju wygryza wąski (1-2 mm) kanalik, żywiąc się płynną zawartością liścia (stadium L1 - plasmophaga). Następnie larwa zaczyna żerować tworząc minę, w fazie początkowej ma ona regularny okrągły kształt o ciemnobrunatnym zabarwieniu i średnicy ok. 5 mm. W okresie późniejszym mina przybiera kształt nieregularny, o zabarwieniu jasnozielonym, osiągając często powierzchnię kilku cm2. Minę tę tworzą kolejne stadia larwalne (L2-L4 - histophaga). Opisane stadia są formami żerującymi; ich ciało jest płaskie, a prognatyczna głowa ma charakterystyczny klinowaty kształt zakończony gryzącym aparatem gębowym. Ostatnie stadium żerujące (L4) przechodzi metamorfozę i zmienia się w larwę cylindrycznego kształtu z głową zaopatrzoną w organy przędne. Zadaniem jej jest wytworzenie kokonu poczwarkowego. Larwa ta także przechodzi wylinkę i powstaje następne stadium larwy przędzącej. Larwy (L5, L6 - aphaga) nie pobierają pokarmu i tkają dwa rodzaje kokonów, letnie (L5) w których poczwarki rozwiną się w tym samym roku i zasilą następne pokolenie motyli, oraz zimowe diapauzujące (L6), z których motyle wylecą w roku następnym. Kokony wykazują zróżnicowanie morfologiczne zależne od pokolenia: letnie są cienkie, koloru białego, natomiast diapauzujące są grube, gęsto utkane, w kolorze brunatno-żółtym. W każdym pokoleniu w liściach zostaje pewien zapas poczwarek diapauzujących, których ilość określa się na 20 do 30% populacji. Stwierdzono, że najwięcej diapauzujących poczwarek występuje w pierwszej wiosennej generacji szrotówka. Poczwarka jest koloru brązowego o długości ok. 3-4 mm; na głowie jej znajduje się kolec, dzięki któremu przebija kokon i blaszkę liściową, poczym motyl może wydostać się na zewnątrz. W minie z reguły zostaje wystająca na zewnątrz pusta osłonka poczwarkowa tzw. exuvium. U poczwarek występuje dymorfizm płciowy (Freise i Heitland 1999): poczwarki samcze różnią się od samiczych wyraźnie szerszym 7 dystalnym segmentem odwłokowym. Warunki klimatyczne środkowej Europy umożliwiają rozwój trzech pełnych pokoleń szrotówka kasztanowcowiaczka w roku. Pierwsze pokolenie pojawia się na początku kwietnia, drugie w końcu czerwca, a trzecie na początku sierpnia. Pełny cykl rozwoju wynosi od około 6 do 9 tygodni. W okresie letnim na liściach kasztanowca występują praktycznie wszystkie stadia rozwojowe tych motyli. | | | | | | Okoliczności poprzedzające pojawienie się gatunku w Polsce | | Po stwierdzeniu go w Macedonii w 1984 r. (Deschka i Dimić 1986), szrotówek zaczął szybko opanowywać kasztanowce w całej Europie (Šefrová i Laštůvka 2001). Najwcześniej stwierdzono go w 1988 r. w Bułgarii (Pelov i in. 1993), a w 1989 r. pojawił się w Austrii, dokąd prawdopodobnie został introdukowany w celach badawczych (Puchberger 1990, Krehan 1995). Kolejne stwierdzenia dotyczyły Chorwacji (Maceljski i Bertic 1995), Włoch , Niemiec (Butin i Führer 1994, Heitland i Metzger 1997), Węgier (Szaboky 1997) oraz Rumunii, Słowacji (Siviček i in. 1997) i Czech (Liska 1997, Skuhravý 1998). W 1998 wykryto go w Szwajcarii (Kenis i Foster 1998), Belgii (De Prons i Puplesienie 2000) i Holandii (Stigter i in. 2000). W 1999 stwierdzono go w Turcji, a w następnym roku w Grecji (Avtzis i Avtzis 2003) oraz we Francji (Guichard i Augustin 2002). W 2002 roku pojawia się w Danii, na Ukrainie (Akimov i in. 2003) i na wyspach brytyjskich. Dalsza jego inwazja na wschód i zachód Europy trwa, na Ukrainie szybko zbliża się do Morza Czarnego, a we Francji przesuwa się w stronę granicy hiszpańskiej. | | | | | | Ekspansja/inwazja w Polsce, czas i miejsce | | Po raz pierwszy stwierdzony w 1998 roku w Wojsławicach koło Wrocławia (Łabanowski i Soika 1998), w następnych latach opanowuje całą południową część kraju, a w 2003 r. występuje już praktycznie na całym obszarze Polski. | | | | | | Występowanie na terenach chronionych | | Według informacji zebranych przez autorow szrotówek został stwierdzony w latach 2006 i 2007 na kasztanowcach rosnących w otulinach i enklawach kilku parków narodowych, m.in. Białowieskiego PN, Ojcowskiego PN, Świętokrzyskiego PN, Tatrzańskiego PN i PN Gór Stołowych. Niewątpliwie występuje też w większości parków krajobrazowych. | | | | | | Wpływ na ekosystemy i gatunki rodzime | | Nie badano. | | | | | | Szkodliwość, profilaktyka i zwalczanie | | Szrotówek w fazie kulminacyjnej ekspansji powoduje znaczne osłabienie drzew kasztanowca, co wprawdzie nie jest zagrożeniem rodzimych biocenoz, jednakże obniża walory estetyczne alej i parków miejskich. Przypadki całkowitego eliminowania pojedynczych kasztanowców są bardzo rzadkie i zwykle mają miejsce na ulicach miast, gdzie stosuje się chlorki w okresie zimowym. Wówczas sumują się szkody wywoływane żerem szrotówka i zasoleniem podłoża glebowego, gdy usuwany śnieg gromadzony jest wokół pni drzew. Stosowane w wielu miastach lepowanie pni jest działaniem szkodliwym dla biocenozy, gdyż niszczy także bezkręgowce drapieżne (nadrzewne pająki, pluskwiaki, sieciarki, muchówki, niektóre chrząszcze itd.), a tylko w mniejszym stopniu motyle szrotówka. Inne stosowane sposoby to iniekcje pniowe i glebowe insektycydów systemicznych (imidacloprid, chliotianidyna) oraz opryskiwanie koron inhibitorami syntezy chityny jak Dimilin. Wpływ tych środków chemicznych na glebę nie został jednak dostatecznie poznany. Powstanie - być może - możliwość biologicznego ograniczania liczebności szrotówka z wykorzystaniem drapieżników i parazytoidów, a także ewentualnie pułapek feromonowych - nad czym trwają badania (Svatos i in. 1999, Grabenweger i in. 2003, Radeghieri 2004).
Należy dodać, iż bardzo spektakularna inwazja szrotówka spowodowała w naszym kraju duże zaniepokojenie i społeczne zapotrzebowanie na informację w tej dziedzinie, czego wyrazem są częste publikacje w prasie fachowej oraz w poczytnych periodykach (np.: Kowalski i Guzik 2000, Ćwikliński i Ćwiklińska 2001, Dankowska 2001, Kosibowicz 2003a, b, c, d, 2005; Morek 2006, i inne). Uaktywniły się również firmy produkujące opaski lepowe oraz preparaty do iniekcji i uruchomiona została akcja reklamowa w tym zakresie. Jednak na razie najwłaściwszą i najbardziej bezpieczną metodą jest profilaktyka - usuwanie na jesieni spod drzew suchych liści i wywożenie ich oraz palenie lub utylizowanie w inny sposób. | | | | | | Prognoza | | Do końca czerwca 2007 r. w południowej Polsce stan ulistnienia kasztanowców był dobry, a zauważalne miny szrotówka nieliczne. Dopiero od lipca tegoż roku obserwowano żółknięcie brzegów blaszki liściowej i miny, głównie jednak na najniżej wyrastających gałęziach, gdzie uszkodzenia objęły 20-30% powierzchni liści; szczytowe części koron były jednolicie zielone i nie wykazywały widocznych uszkodzeń. Wiosną wszystkie kasztanowce kwitły obficie, a w sierpniu wielkość owoców nie odbiegała od normy sprzed dwóch dekad. Stwierdzony stan może oznaczać początek etapu stabilizacyjnego.
Większe uszkodzenia notuje się nadal w Polsce północnej, aczkolwiek i tam stan ulistnienia kasztanowców jest znacznie lepszy niż w dwóch poprzednich sezonach, można więc określić obecny etap jako początek stadium post-kulminacyjnego. Prawdopodobnie stadium kulminacyjne inwazji przesuwa się obecnie na obszary ościennych państw wschodnio-bałtyckich i Białorusi, tam też w najbliższych latach mogą wystąpić największe uszkodzenia kasztanowców. W ciągu 2-3 lat faza stabilizacji szrotówka powinna nastąpić także w rejonach północnych kraju. Początków fazy regresji można się spodziewać dopiero za kilka lat. Prawdopodobnie podobny przebieg będzie miał ten proces w Niemczech. Natomiast poza wschodnią granicą następstwo faz będzie zapewne przesunięte w czasie o 2-5 lat. | | | | | | Literatura | | - Akimov I., Zerova M., Gershenzon Z., Narolski N., Kokhanets A., Sviridov S. 2003: Pervoe soobshchenie o pojavleni w Ukrainie kashtanovoi miniurushchey moli Cameraria ohridella (Lepidoptera, Gracillariidae) na konskom kashtane obyknovennom Aesculus hippocastanum (Hippocastanaceae). Vestnik zoologii, Kiev 37 (1): 3-12.
- Avtzis N., Avtzis D. 2003. The Attack of Aesculus hippocastanum L. by Cameraria ohridella Deschka and Dimic (Lepidoptera: Gracillariidae) in Grece. Proceedings Ecology, Survey and Managment of Forest Insects. Kraków, Poland, September 1-5, 2002, Published by USDA Forest Service Northeastern Research Station, General Technical Report NE-311: 1-5.
- Butin H., Führer E. 1994: Die Kastanien-Miniermotte (Cameraria ohridella Deschka & Dimic) ein neuer Schädling an Aesculus hippocastanum. Nachrichtenbl. Deut. Pflanzenschutzdienst, 46: 89-91.
- Ćwikliński L., Ćwiklińska J. 2001. Nieco więcej o szrotówku kasztanowcowiaczku. Ochrona Roślin nr 5-6: 10-12.
- Dankowska E. 2001. Nowe zagrożenie dla kasztanowców. Szrotówek kasztanowcowiaczek (Cameraria ohridella). Aura 6: .
- De Prins W., Puplesiene J. 2000. Cameraria ohridella, een nieuw soort voor de Belgische fauna (Lepidoptera: Gracillariidae). Phegea 28 (1): 1-6.
- Deschka G., Dimic N. 1986. Cameraria ohridella n. sp. aus Mazedonien, Jugoslawien (Lepidoptera, Lithocelletidae). Acta Entomologica Jugoslavica 22 (1): 11-23.
- Freise J., Heitland W. 1999. A brief note on sexual differences in pupae of the horse-chestnut leafminer, Cameraria ohridella Deschka & Dimic (1986) (Lepidoptera, Gracillariidae), a new pest in Central Europe attacking Aesculus hippocastanum. Journal of Applied Entomology 123: 191-192.
- Freise J., Heitland W. 2001. Neue Aspekte zur Biologie und Ökologie der Roßkastanien-Miniermotte, Cameraria ohridella Deschka & Dimic (1986) (Lep., Gracillariidae), einem neuartigen Schädling an Aesculus hippocastanum. Mitt. Dtsch. Ges. Allg. Angew. Ent. 13: 135-139.
- Grabenweger G., Stolz M., Jeziorny K. 2003. A key to the parasitoids of Cameraria ohridella (Lepidoptera, Gracillariidae). Institute of Plant Protection University of Natural resources and Applied Life Sciences. Wien, Austria: .
- Gregor F., Laštůvka Z., Mrkva R. 1998. Horse chestnut leafminer also found on maple. Ochrona-Roślin 34 (2): 67-68.
- Guichard S., Augustin S. 2002. Acute spread in France of an invasive pest, the horse chestnut leafminerCameraria ohridella Deschka & Dimic (Lep., Gracillariidae). Journal of Pest Science 75: 145-149.
- Heitland W., Metzger J. 1997. Die Kastanienminiermotte Cameraria ohridella Deschka & Dimic (Lep., Gracillariidae) in Bayern. LWF Aktuell, Juni 1997: 16-17.
- Kenis M., Foster B. 1998. Die Rosskastanien-Miniermotte: neu in der Schweiz. Gartenbau 39: 16-17.
- Kosibowicz M. 2003a. Metody ograniczania populacji szrotówka kasztanowcowiaczka. Las Polski 17: 12-13.
- Kosibowicz M. 2003b. Szrotówek atakuje jawory. Las Polski 17: 13
- Kosibowicz M. 2003c. Metody ograniczania populacji szrotówka kasztanowcowiaczka. Las Polski 17: 12-13.
- Kosibowicz M. 2003d. Nad nimi krąży motylek. Strona, Kraków 8 (17): 22-23.
- Kosibowicz M. 2005. Szrotówek kasztanowcowiaczek Cameraria ohridella Deschka & Dimić (Lepidoptera, Gracillaridae), nowy inwazyjny szkodnik kasztanowca białego Aesculus hippocastanum L. w Polsce - biologia i metody zwalczania. Leśne Prace Badawcze 2: 121-132.
- Kowalski T. 1999. Nowy szkodnik liści kasztanowca. Trybuna Leśnika nr 10: .
- Kowalski T. Guzik G. 2000. Pojawienie się szrotówka na kasztanowcu. Głos Lasu nr 9: .
- Krehan H. 1995. The horsechestnut leafmining moth Cameraria ohridella - incidence of attack in Austria. Forstschutz Aktuell No. 16: 8-11.
- Liska J. 1997. Verbreitung der Roßkastanienminiermotte in der Tschechischen Repubklik. Forstschutz Aktuell, 21: 5.
- Łabanowski G., Soika G. 1998. Cameraria ohridella damages horse chestnut trees in Poland. Ochrona Roślin 42: 12.
- Maceljski M., Bertic D. 1995. Kestenov moljac miner - Cameraria ohridella Deschka & Dimic (Lepidoptera, Gracillariidae) - novi opasni stetnik u Hrvatskoj. Fragmenta phitomedica et herbologica, 23 (2): 9-18.
- Morek G. 2006. Szrotówek kontratakuje. Polityka nr 17/18: 152-153.
- Pelov V., Tomov R., Trenchev G. 1993. Cameraria ohridella (Deschka & Dimic (Gracillariidae, Lepidoptera) - neuer Schädling an Roßkastanien (Aesculus hippocastanum L.) in Bulgarien. Natural Sciences Forest Protection Conference, Sofia: 95-98.
- Pschorn-Walcher H. 1997. Zur Biology und Populationsentwicklung der eingescleppten Rosskastanien-Minermotte, Cameraria ohridella. Forstschutz Aktuell. No 21: 7-10.
- Puchberger K.M. 1990. Cameraria ohridella Deschka & Dimic (Lep., Lithocolletidae) in Oberösterreich, Steyrer Entomologische Runde 24: 79-81.
- Radeghieri P. 2004. Cameraria ohridella (Lepidoptera, Gracillariidae) predation by Crematogaster scutellaria (Hymenoptera: Formicidae) in Northern Italy (Preliminary note). Bulletin of Insectology 57 (1): 63-64.
- Šefrová H., Laštůvka Z. 2001. Dispersal of the horse-chestnut leafminer, Cameraria ohridella Deschka & Dimic, 1986, in Europe: its course, ways and causes (Lepidoptera: Gracillariidae). Entomologische Zeitschrift 111 (7): 194-198.
- Siviček P., Hrubik P., Juhasová G. 1997.Verbreitung der Roßkastanienminermotte in der Slovakei. Forstschutz- Aktuell, Wien 21: 6.
- Skuhravý V. 1998. Zur Kenntnis der Blattminen-Motte Cameraria ohridella Desch. & Dim. (Lep., Lithocolletidae) an Aesculus hippocsatanum L. in der Tschechischen Republik. Anzeiger für Schädlingskunde Pflanzenschutz Umweltschutz 71: 82-84.
- Skuhravý V. 1999. An overview of knowledge about the horsechestnut miner Cameraria ohridella Desch. & Dim. (Lep., Gracillariidae). Anz. Schädlingsk. 72 (4): 95-99.
- Stigter H., Van Frankenhuyzen A., Moraal L.G. 2000. De paardenkastanjemineermot, Cameraria ohridella, een nieuwe bladmineerder voor Nederland (Lepidoptera: Gracillariidae). Ent. Ber. Amst. 60 (8): 159-163.
- Svatos A., Kalinová B., Hoskovec M., Hovorka O., Hrdy I. 1999. Identification of a new lepidopteran sex pheromone in picogram quantities using an antennal biodetector: (8E,10Z)-Tetradeca-8,10-dienal from Cameraria ohridella. Tetrahedron Letters, 40 (38): 7011-7014.
- Szabóky C. 1997. Verbreitung der Roßkastanienminiermotte in Ungarn. Forstschutz Aktuell, Wien 21: 4.
| | | | | | Streszczenie | | Horse Chestnut Leafminer Cameraria ohridella Descha & Dimić is a small moth, a monophage of horse-chestnut Aesculus hippocastanum or an oligophage of the genus Aesculus. It was described from Macedonia, already as a strongly invasive species, noted for the first time in 1984. Its origin is uncertain; it could be that a native element from the borderland of Balkan Peninsula and Asia Minor (the natural range of Ae. hippocastanum), which had escaped detection by enthomologists, became suddenly active, or that the species arrived by air transport from remote areas, the Neoarctic origin having been most probable. From Macedonia Horse Chestnut Leafminer spread during a dozen of years all over the whole Balkan Peninsula and Central Europe. In Poland it was first noted in Lower Silesia (environs of Wrocław) in 1998, and five years later it occurred in the whole country. Imagines have poor ability to fly, and only at short distances. However, if assisted by wind they can move at large distances (even to several tens of kilometers). Females lay eggs on the upper side of leaves and hatching larvae mine leaf blade where they next pupate in special cocoons. In one year three or four generations can be produced and some of larvae in each generation produce harder cocoons in which they overwinter. This phenomenon allows one to apply the most effective preventive measures: raking up, removing and destroying fallen leaves. In this way one can reduce to the minimum the numbers of spring generation. Expensive covering the horse chestnut trunks with sticky traps, which was until quite lately applied in many towns in spring did not meet expectations, what more, destroyed its potential natural enemies (spiders, predatory insects etc.). During last years in the south of Poland horse chestnut trees are healthy up to the end of June, and next one can observe damage caused by summer generations of Horse Chestnut Leafminer, brought most probably by wind from other places. In areas when the activity of the pest is declining one can observe serious damage to the foliage of the lowest branches, while leaves from the higher parts of crowns show only a small number of mines. It is observed that trees growing along main streets are more damaged because these trees are additionally weakened by winter salting of roadways. In northern Poland the percentage of damaged trees is much higher which indicates that the peak phase of invasion continues in this region. | | | | | | Opracowanie | | Mieczysław Kosibowicz i Jerzy Pawłowski | | | | |
|
)
)